|
NATIONAL PARK SERVICE
The Grizzlies of Mount McKinley |

|
CHAPTER 2:
Study Background
Classification and Characteristics
Taxonomy of Grizzly and Brown Bears
C. Hart Merriam (1918) published a classification of the grizzly and brown bears. For several years he had been gathering material, chiefly skulls, from hunters and others. He recognized 86 forms, most of them full species. Mammalogists early questioned the validity of the many species, most of which Merriam himself described. Judgments in the field of taxonomy are questioned frequently but usually not to the extent that the grizzly species were. It was believed that Merriam's entire grizzly classification was based on false premises, that he had assumed wrongly that variation found in skulls represented species rather than types of individual variation, such as we find in humans.
In the early 1930s my brother Olaus and I had the pleasure of spending an evening with Dr. Merriam in his Washington, D.C., home. He was a most colorful individual and an outstanding raconteur. He regaled us with stories of his biological explorations in the West, from as far back as the 1870s. He also told of an incident that concerned the dispute surrounding grizzly taxonomy and Theodore Roosevelt, who was among the doubters of the many grizzly species recognized. The incident took place at the Cosmos Club during a meeting to which President Roosevelt was invited. Dr. Merriam arrived early, carrying two grizzly skulls which he placed on the mantelpiece. Roosevelt soon spied the skulls, and a lively discussion followed. He conceded that the skulls were different enough to represent two species. Then followed Dr. Merriam's triumph—he told the President that the skulls represented two species which he had questioned. Merriam thus clinched his argument about his grizzly taxonomy and no doubt added to his certainty concerning the validity of his many species. My brother and I were sorry that we were still among the doubters.
Robert Rausch (1953), who has discussed bear taxonomy in Alaska, found wide variations in a number of skulls (22 with full data) gathered in the Brooks Range. He concluded that he was dealing with individual variation in an interbreeding bear population. He and others have, in recent years, placed the North American grizzlies and brown bears, along with the Eurasian brown bears, in a single species, Ursus arctos. Rausch lumps about 14 species listed for northern and central Alaska into one subspecies, Ursus arctos horribilis. Following Rausch's classification, the grizzly in McKinley National Park would be called Ursus arctos horribilis.
Description
In the field a grizzly bear may be distinguished readily from the black bear because of its pronounced shoulder hump. Also its facial profile differs from the straight profile of the black bear in being somewhat dished, that is, the forehead tends to rise and give a break in the profile line (Fig. 2). Black bears are generally black or brown, while grizzlies show a wide range of intermediate hues and have a grizzling over much of the pelage.

|
| Fig. 2. The shoulder hump and dished facial profile easily distinguish grizzlies from black bears in the field. |
In contrast to black bears, which have rather short and very curved claws on the forefeet, grizzlies have very long claws on the forefeet, more than 2 inches long unless badly worn, and only slightly curved. The middle foreclaws of an adult female measured 3-1/2 inches along the dorsal curve and 2-1/2 inches in a straight line from the base to the tip. Digging for roots and for ground squirrels tends to wear away the tips. The claws on the hind feet are much shorter and curved more sharply. The color of the claws in grizzlies can vary from dark brown to almost white (Figs. 3, 4)
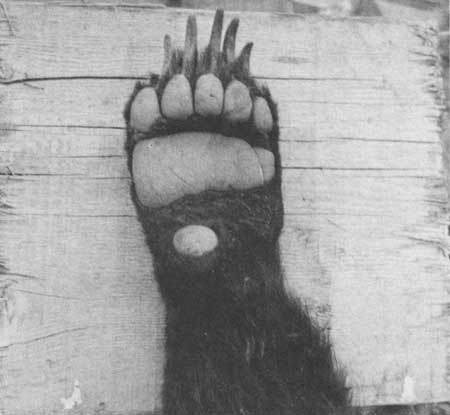 
|
| Figs. 3,4. Forefeet (top) and hind feet (bottom) of grizzlies are clearly distinguishable. The long claws on the forefeet show in most tracks and contrast with the shorter, more curved claws of black bears. |
The relatively straight foreclaws are not suited for climbing tree trunks. I have seen spring cubs and yearlings climb 10 or 12 feet from the ground in willows, clambering about in play. Ordinarily, adult grizzlies do not engage in tree climbing even in areas where trees are plentiful.
Grizzly bears have up to 42 teeth. One or more of the small premolars may be missing. The molar teeth are considerably flattened, apparently an adaptation to the omnivorous diet. However, the mastication of the vegetation eaten is very slight, as is shown by the remains found in the droppings.
The majority of the grizzlies in the park are light tan over most of the body, often referred to as blond. Head, neck, and shoulders may be light, almost buffy white, with legs and belly dark brown (in fall fur is shorter and darker.) Some are reddish-brown, some a rich dark chocolate, and a few almost black. An old male I examined was black except for dark-brown grizzling over the shoulders and back. The feet, legs, and underparts of the body are dark in all color phases, varying from blackish to various shades of dark reddish-brown. The face is generally slightly darker than the body but an occasional bear has such a light face that the dark eyes show prominently. The pelage is usually grizzled, the tips of the hairs being light. This is not evident in many dark bears. The shade of color of a bear varies according to the direction from which the light strikes it, relative to the position of the viewer. The bear appears darker when facing away from the light because of reduced reflection. On one occasion a tourist told me that he had seen a bear on Sable Pass that was light on one side and black on the other. I had noticed this striking difference in this particular bear too. Both sides were alike in color but as the bear shifted its position in relation to the light, the color tended to vary from blondish to blackish. The effect of light direction on the color of this blondish bear was more extreme than noted in any other bear. Possibly this was due to some special character of the grizzling in this individual.
The large grizzlies of the Alaska coast and adjacent islands, generally called brown bears, are colored more uniformly than bears in the interior, and usually are dark brown, although occasionally light-colored ones are reported in coastal populations.
Pelage color usually undergoes seasonal change. The grizzlies in McKinley National Park emerge from hibernation with their autumn coats still in excellent condition. As spring progresses, the fur tends to fade and lighten in color. The long northern days and light reflection from snowfields probably accelerates this fading. By July the fur on a few bears becomes somewhat ragged in a patch or two, but in most bears the changing of coat is hardly perceptible.
In my notes over the years I find references to indications of shedding in some bears. These observations are concentrated in the month of July, some in August, and one year I noted the shedding of a 4-year-old was still not completed on 18 September. Usually by September the bears, with few exceptions, have new coats. They are much darker than the old coats and appear rich and alive.
The spring cubs are blackish or dark brown in their spring coats. Some have a white vertical streak on the sides of the neck which usually is lost by the time they are 2 years old. By August they have acquired a new coat and show a lighter grizzling that is more pronounced in those cubs that were brownish in the spring.
Little information is available on weights of grizzlies in McKinley National Park. An old male grizzly that was not very fat weighed 650 pounds. The animal had been shot so there had been a loss of blood which could not be calculated. There was no opportunity to weigh a female but it is likely that females weigh about 200 pounds less than males. The difference in size shows up strikingly when one observes a mated pair.
The measurements of an old male and an old female from the park are as follows:
| Sex | Date | Total length inches |
Tail inches |
Hind foot length inches |
Front foot width inches |
Weight pounds |
| Male | 11 Sept. 1951 | 73 | 7 | 11 | 6.4 | 650 |
| Female | 28 Aug. 1963 | 56 | 9 | 5.25 |
The difference in size between the sexes also is indicated clearly in skull measurements. A male skull, 15.8 inches long, weighed 4 pounds 12 ounces; a female skull, 13 inches long, weighed 2 pounds 7 ounces.
In the Brooks Range, Dr. Robert Rausch found grizzlies weighing from 400 to 700 pounds. The range in size of adult bears in the park is apparently similar to that from the above locality. The coastal and island grizzlies are much larger and are known to attain a weight of at least 1,200 pounds. Estimates run even higher. It is thought that coastal bears attain their large size because of the abundance of protein foods, mostly fish.
Age and Mortality
In zoos, grizzlies have lived for almost 30 years. That some bears in the wild live to a ripe old age is shown by the thorough wear we find on the teeth. One old male grizzly showed excessive tooth wear: four of the molariform teeth were worn in two, only two root stubs remaining in each, and one molar was missing; the upper and lower incisors were worn to the gums; the two upper canines were worn but still retained their shape, but the two lower canines were worn until only blunt stubs remained. This old male, shot as he was breaking into a work camp after several raids, had lived long enough to have worn out his teeth.
A female, mother of three spring cubs, and apparently killed by another bear, was quite old. The teeth were not as worn as those of the male described above, but the incisors and molariform teeth were worn to the gums. The canines showed much wear and were quite blunted.
Because grizzlies are relatively scarce, it is seldom that one finds bone remains or a carcass, so little was learned of relative mortality rates in different age groups. Nevertheless, it may be of interest to list skull remains and carcasses found in the field.
Skull remains:
1. Mandible of spring cub.
2. Mandible of what appears to be a yearling.
3. Part of skull of young adult bear.
4. Skull of old female with some necrosis at base of a lower molar.
Carcasses found:
1. Two spring cubs killed by another female.
2. Very old female, mother of three spring cubs, at garbage dump. She was killed apparently by another bear.
3. A young adult female apparently killed by another grizzly.
4. Adult bear—cause of death not known.
From what little evidence is available, it appears that death at the hands of another grizzly may be a large part of mortality. Sufficient information to assess the effects of disease and parasites is lacking.
Use of Senses
Like many other mammals, grizzlies rely extensively on their sense of smell in conducting their day-to-day activities, although sight and hearing also play a role. Initial awareness of the presence of potential prey, other bears, or possible competition for carrion such as wolves or wolverines usually seems to be accomplished by detecting their scent.
I have watched bears, nose to ground, move about as though following a trail in areas being traversed by migrating caribou. Once a bear behaved in this manner for several minutes, eventually flushing out a female caribou with a young calf and successfully capturing the calf. It seemed to know by the scent that a caribou calf was in the area.
On some occasions when a grizzly is concentrating on ground squirrels, it may hear one calling or see it, gallop to the spot where the squirrel enters a burrow, and begin to dig it out. The squirrel sometimes escapes from another exit, unnoticed, while the bear concentrates on digging. Although the escape was not observed, the bear soon ceases in his efforts, realizing, by the lack of ground squirrel odor presumably, that his quarry has left. Then he either moves on or follows the trail of the squirrel to another burrow and exerts himself again. In one instance, a bear followed the escapee's trail for about 100 feet and was rewarded for the effort by capturing the unlucky squirrel. More often, an escaping ground squirrel is seen by the bear and pursued immediately to the next burrow.
Bears are reported to have relatively poor vision, at least at long distances. My observations do not contradict this. Individual visual recognition, within families for instance, sometimes appears to be unreliable, especially if cubs become separated from mothers by several hundred yards. At such times there can be much hesitation on the part of the cub to rejoin the mother, even though they are in sight of each other. Olfactory reassurance that an adult is indeed its mother seems prerequisite for a cub to resume its usual activity within the family.
The use of hearing by bears is not as obvious as that of sight and smell. Cubs do respond at some distance to low "woofs" or grunts from the mother; and I have noted that bears can detect slight noises that I have made at distances of 200 yards. Even though it may not play a prominent role in their activities, I believe grizzlies do have an acute sense of hearing.
Habitat
Much of McKinley National Park is treeless tundra, but strips of woods follow the rivers far into the park, and patches of trees grow here and there on adjacent mountain slopes. Timberline varies according to soil and exposure; in places it reaches elevations of over 3,500 feet.
White spruce is the common conifer. Black spruce is confined to poorly drained and boggy areas. Along the north boundary I have seen a few patches of tamarack. Cottonwood and aspen are distributed widely and a few birches grow at lower elevations. Along the McKinley River an extensive strip of cottonwoods may be seen from the highway.
The tundra supports a growth of willow and dwarf birch. Over 20 kinds of willow occur in the park. They range in size from small forms only 2 or 3 inches in height to brushy growths 20 feet tall. In places, the small willows may grow dense enough to form a sod. These shrubs are highly important for wildlife. Alder brush is distributed widely and is plentiful on canyon slopes; near Wonder Lake there are many clumps of alder in the rolling tundra.
The low ground cover over the park consists of mosses, lichens, sedges, grasses, horsetails, and herbaceous plants—many species of each. Early flowers may begin to bloom in late April and early May, and at the higher elevations some blooms may be seen in later summer.
All of McKinley National Park can be considered bear country, except for the snow-covered upper reaches of the peaks of the Alaska Range. One may meet grizzlies anywhere, from river bars to ridge tops. Particular habitats used by bears vary with the season and from year to year, depending on food availability. In the spring, the river bars and some hillsides are favored places for digging roots of peavine and other plants. As green vegetation becomes available, many bears move to areas with grassy swales such as on Sable Pass where grazing, primarily on grass, becomes a principal activity. Berry crops appear later in the summer and the location of bear activity coincides with areas where blueberries, crowberries, or buffaloberries are abundant. In some years when berry crops are poor, bears wander more widely than usual in late summer. The habitat is sufficiently varied over most of the park that bears may find spring, summer, and autumn foods within a limited area.
Thus, bear habitat is affected by the vagaries of weather and its effects on the phenology of plant foods of bears. Another direct influence on bear habitat, though one that operates over the long term, is the change wrought by rivers on river bars. The many rivers in the park, such as the Sanctuary, Teklanika, East Fork, and Toklat, break up into numerous channels which are constantly shifting over the broad gravel bars. During the summer, they carry much glacial silt that is picked up in one place and deposited in another. Where the silt is deposited, the bottom of the channel may build up until the stream breaks over the edge to flow off to one side over a slightly lower part of the bar. Heavy rains greatly increase the volume, sometimes forming one sheet of water covering the entire river bed and causing many changes in the channels. By moving back and forth, the streams tend to keep widening the bars. Some bars are a half-mile or more in width (Fig. 5). Changes may be slow or rapid.

|
| Fig. 5. Here a glacial stream has shifted its channel and is washing away an old vegetated river bar where bears, spring and fall, feed on roots. Old river bars are continually washing away and new ones forming. |
Over long stretches of the rivers, gravel bars have remained undisturbed long enough to become covered with a thin, firm sod. These old bars are delightful for hiking as they are covered chiefly with low-growing vegetation and are as smooth as a lawn. The grizzlies also find them delightful and spend much time in spring and fall digging the roots of the peavine which prospers in this habitat. Up toward the heads of the rivers another species of the pea family (Oxytropis viscida) flourishes and attracts the grizzlies who come to graze on its stems, leaves, and flowers. Some of the old bars support good stands of buffaloberry, and these berries are an important food for bears. Thus it is apparent that the river bars represent a significant part of bear habitat.
These old river bars originate through the activity of the rivers, but the river may also destroy them. An old bar that has been left in repose for 50 or more years may suddenly be invaded by a shift of some of the stream branches. In the mid-1950s, part of the Toklat River swung westward and flowed over an extensive old bar, cutting new channels which braided and widened. This had been a favorite rooting area for the grizzlies. Similarly, along the East Fork River a channel swung sharply into an old, high river bar, supporting a good stand of buffaloberry, and reduced the size of this bar considerably.
In places, river bars may be left undisturbed long enough for choice grizzly habitat to disappear because of plant succession. Along the Teklanika River an old river bar has supported a good stand of peavine for years. Now a young spruce forest is taking over and is slowly causing a decrease in the peavine. In time, the peavine will be shaded out entirely unless the stream invades.
The duration of the cycle involving the change from gravel bar to vegetation-covered bar and back to gravel bar varies greatly. In some places, plant life is washed away before it has had time to form a good sod. In other, limited areas, a spruce forest has had time to develop before the stream has begun to erode, invade, and return the wooded area to a barren gravel bar (Fig. 6). Thus the cycle tends to keep the grizzly habitat along the rivers in balance.

|
| Fig. 6. Here the glacial stream is washing away a wooded flat, thus extending the width of the bar and creating more peavine habitat for bears in the future. |
Other habitats in the park frequented by grizzlies are not subject to changes of this sort. The progress of plant succession in some areas is very gradual in this northern climate, and long-term weathering processes occur, but most parts of the bear's domain are not altered drastically over the years.
Numbers and Density
Total numbers of bears in the park and changes from year to year are difficult to determine. Several factors contribute to this difficulty. One is size of area and rugged topography which precludes consistent sightings of bears, even in portions of the park visible from the road. The majority of effort was in areas visible from the road; year-to-year variations in season and relative abundance of various food sources resulted in different patterns of use by bears. Thus, in some years bears were concentrated in areas where sightings were relatively easy, whereas in other years bears were absent from these areas, resulting in fewer sightings. In addition, my intensity of effort varied from year to year, and prior to about 1959 there was no special concentration on grizzlies.
Few aerial counts of bears have been made in the park, but aerial counts probably are not as complete as ground counts anyway. In 1969, a pilot surveyed the park for bears and remarked on the scarcity of females with cubs; he saw mainly lone bears. This is the year in which I recorded 20 families, more than in any other year. Other bear researchers have remarked on the difficulty of spotting grizzlies from the air even when a bear has a radio collar and its general location is known (Herrero 1972:82).
For each year I have calculated minimum numbers of different bears seen. These figures are probably fairly accurate for families because variations in pelage characteristics of the cubs and females make individual identification possible in most cases. Numbers of lone bears, however, probably are substantially underestimated. I have calculated numbers conservatively by counting only those lone bears and families that I am confident are different from other sightings. These data are presented in Table 1.
Table 1. Minimum numbers of grizzly bears observed in Mt. McKinley National Park,
1939-1970.
| Year | Size of litter | Females with cubs | Lone bears |
Total adult bears | Total bears | ||
| 1 | 2 | 3 | |||||
| 1970 | 7 | 4 | 0 | 11 | 10 | 21 | 36 |
| 1969 | 12 | 8 | 0 | 20 | 18 | 38 | 66 |
| 1967 | 5 | 6 | 2 | 13 | 17 | 30 | 53 |
| 1966 | 5 | 4 | 1 | 10 | 15 | 25 | 41 |
| 1965 | 3 | 5 | 0 | 8 | 15 | 23 | 36 |
| 1964 | 5 | 8 | 0 | 13 | 28 | 41 | 62 |
| 1963 | 9 | 9 | 1 | 19 | 24 | 43 | 73 |
| 1962 | 6 | 9 | 0 | 15 | 29 | 44 | 68 |
| 1961 | 5 | 10 | 0 | 15 | 36 | 51 | 76 |
| 1960 | 4 | 13 | 0 | 17 | 22 | 39 | 69 |
| 1959 | 5 | 13 | 1 | 19 | 23 | 42 | 77 |
| 1956 | 5 | 6 | 1 | 12 | 19 | 31 | 51 |
| 1955 | 4 | 6 | 1 | 1l | 17 | 28 | 47 |
| 1953 | 3 | 2 | 2 | 7 | 13 | 20 | 33 |
| 1951 | 2 | 5 | 0 | 7 | 7 | 14 | 26 |
| 1950 | 0 | 1 | 1 | 2 | 11 | 13 | 18 |
| 1949 | 2 | 6 | 1 | 9 | 17 | 26 | 43 |
| 1948 | 3 | 5 | 1 | 9 | 12 | 21 | 37 |
| 1947 | 4 | 5 | 0 | 9 | 17 | 26 | 40 |
| 1945 | 1 | 0 | 0 | 1 | 9 | 10 | 11 |
| 1941 | 2 | 1 | 2 | 5 | 11 | 16 | 26 |
| 1940 | 1 | 3 | 3 | 7 | 14 | 21 | 37 |
| 1939 | 4 | 3 | 2 | 9 | 13 | 22 | 38 |
Numbers of different bears seen were particularly low in 1965, 1966, and 1970. There was no apparent reason for low numbers recorded in 1965. In 1966, spring was very late in the park and bears were not abundant in areas usually favored by them in other years. The information from 1970 is incomplete because I was in the park only from late May through June.
Local densities of bears within the park are difficult to calculate because bears move around considerably during the summer as food habits and food availability change. Greatest densities occur on Sable Pass. In 1961 and 1962, there were as many as 3-1/2 to 4-1/2 bears per square mile. A rough figure of density for the portion of the park that my observations covered can be arrived at by using an area 5 miles wide along the length of the road from park headquarters to Wonder Lake, a distance of about 80 miles. Densities in this 400 square mile area in the years from 1959 to 1970 ranged from 0.9 to 1.9 bears per 10 square miles or, if cubs are omitted and only lone bears and family units are used, 0.5 to 1.3 bears per 10 square miles.
Pearson (1972) estimates a density of one grizzly per 10 square miles in Yukon Territory and Kistchinski (1972) suggests densities in northeast Siberia of from 1-1/2 to 2-1/2 bears per 10 square miles. In Glacier National Park, Montana, Martinka (1974) estimated a density of 1 grizzly per 8.2 square miles. Densities may be much higher, comparable to those recorded on Sable Pass, in small areas on the southwest coast of Alaska and in northeast Siberia where bears congregate to take advantage of a particular food source. Thus, density of grizzly bears in McKinley National Park is not dissimilar to that in other areas (Fig. 7)

|
| Fig. 7. A female grizzly with two spring cubs on Sable Pass, an area of the park favored by bears in summer for grazing. |
My information on production of young also shows low production in the same 3 years that numbers were low, probably a result of bears being less observable in those years rather than actual lower recruitment rates. There seems to be no strong relationship between number of spring litters and number of litters of older cubs. Additional information on family statistics and breeding interval is presented later.
Age Determination of Cubs
In these McKinley National Park studies, I have separated cubs into three categories: first year or spring cubs, yearlings, and 2-year olds. In a few instances, cubs 3 and 4 years old were recognized because of my earlier acquaintance with them.
Spring Cubs
Data from bears living in zoos indicate that cubs are born in January and February after a gestation period of about 7 months. Seton (1929) writes that a newborn grizzly was only 8-1/2 inches long, had a grizzly shoulder hump, a tail proportionately longer than that of an adult, and weighed 1-1/2 pounds. It appeared to be naked but was covered with fine, short gray hair. In 1 hour and 40 minutes it began to nurse a foster-mother dog.
When the tiny cubs are observed abroad in spring and early summer, they still are surprisingly small and scrawny. They seem too tiny to be bears (Fig. 8). Their color is blackish, but on close look some are dark brown. These latter apparently become lighter, blondish bears as they age. Some cubs have a white vertical streak on one or both sides of the neck. The amount of white may vary from a thin line to a rather extensive patch, and can be used to identify the individual reliably (Fig. 9). Growth of cubs during the summer is slow. They become more roly-poly, and the fur be comes grizzled by fall. There is no difficulty in recognizing the spring cubs throughout the summer.

|
| Fig. 8. Mother with spring cub. The whitish strip on the cub's neck still shows in September. |

|
| Fig. 9. Some spring cubs have distinct white patches on the sides of the neck which usually disappear by the time they are 2 years old. |
Yearlings
The yearlings in spring are about the size of the spring cubs in autumn. They are obviously not spring cubs and are too small for 2-year-old cubs, although even an experienced hunter may confuse them at times. Judging from field observations, there is sometimes considerable variation in the size of cubs in different litters in the same age categories. I recall a family of two yearlings that I knew as spring cubs that were especially small. A rather experienced bear-hunting guide thought they were spring cubs. When these cubs were 2 years old, they were still small and seemed too small for 2-year-old cubs. But their age was always determinable. A variation in size of cubs is sometimes shown strikingly in a single litter where one cub may be much larger than the other. Toward autumn, yearlings seem to be the size of spring 2-year-old cubs. At this time one might wonder occasionally whether cubs are yearlings or 2-year-olds if the yearlings happen to be especially large (Fig. 10).

|
| Fig. 10. Mother with yearling (same family as Fig. 8) in spring of 1964. |
Two-Year-Olds
Two-year-old cubs show considerable variation in size. I have seen some suckling that seemed too large for this age. Perhaps they were males and their birthdays fell in the early part of the parturition period. Small two-year-olds, if still with the mother by late summer, could perhaps seem small enough for large yearlings if one were not familiar with them. Families in these categories may be somewhat puzzling to the observer when seen for the first time (Fig. 11).

|
| Fig. 11. Mother with 2-year-old cubs feeding on first green grass blades (June 23, 1963). |
In the field, I frequently have noted that the size of cubs compared to their mother seems to vary a great deal from day to day. At times the cubs seem large, then again, small. I also have noted in examining several pictures of a family that the cubs seem large in some pictures and small in others. On the whole, with experience in observation, one usually can be quite sure of age determinations in the three age categories described here. On five occasions I saw a 3-year-old cub in the spring still with its mother. If I had not known the family, I would have assumed the cub to be a 2-year-old. After seeing this cub still with its mother, it occurred to me that on one or two other occasions cubs were 3-year-olds rather than 2-year-olds as I had assumed.
After the cubs have left their mothers, size is difficult to determine because there is no good method of comparison. A 2- or 3-year-old cub seen alone might be taken for an older bear. The body seems shorter in young bears but this is an uncertain criterion. Once I saw two 2-year-old cubs near other older cubs and could easily recognize them as smaller. But when I saw these two by themselves, their smaller size was not obvious.
It has always seemed to me that the closer one approaches a bear, especially a cub, the smaller he appears to be. (If danger is involved, of course, the opposite may be true.) I have been close to known 2-year-old cubs that seemed to be the size of yearlings. On one occasion, the remarks of a friend of mine were significant in regard to judgment of size. The mother of three spring cubs that had been visiting a garbage dump was found dead. When my friend, who had often seen the mother alive, saw the carcass, he exclaimed that this could not be the mother of the three cubs he knew because that mother was "huge" and this dead one was small. Yet her dark color and the presence of the orphans made identification certain. Incidentally, several of the photographers in the park became quite expert in recognizing ages of cubs and also in recognizing the different families. But, as a final word, size in big country is always deceptive.
Some Family Statistics
The usual number of cubs in a litter varies from one to three. An old timer reported that he once saw a litter of four cubs in McKinley National Park; in other areas also, four cubs have been reported and litters of four have been recorded in zoos. Over a period of years I have recorded the number of cubs in 249 families for most of which the ages were known (Table 2).
Table 2. Frequency of litter sizes at different ages observed in grizzly bears in Mt. McKinley National Park.
| 1-cub-family | 2-cub family | 3-cub family | Totals | Mean litter size | |
| Spring cubs | 21 | 36 | 11 | 68 | 1.85 |
| Yearlings | 25 | 41 | 5 | 71 | 1.72 |
| Two-year-olds | 30 | 38 | 1 | 69 | 1.58 |
| Three-year-olds | 5 | 1 | 0 | 6 | 1.14 |
| Unknown age | 14 | 19 | 2 | 35 | 1.66 |
| Total | 95 | 135 | 19 | 249 | 1.70 |
In a study of the Kodiak bears at a salmon stream, Troyer and Hensel (1964) found that 51% of 39 spring-cub litters contained three cubs, 26%, 2 cubs, and 23%, 1 cub. The mean size of the spring-cub litters of these bears was 2.36. In McKinley Park, the mean litter size of the spring cubs was only 1.85. The high protein fish diet of the Kodiak bears possibly accounts in part for the large litters, and the longer season and more favorable climate also may be factors. In other studies of grizzlies, mean litter sizes have been reported as follows: 2.12 in Glacier National Park, British Columbia (Mundy 1963); 2.19 in the Alaska Peninsula (Lentfer 1966); 1.58 in Kluane National Park, Yukon Territory (Pearson 1972); 2.2 in Yellowstone National Park (Craighead and Craighead 1967); and 1.7 in Glacier National Park, Montana (Martinka 1974).
Table 3 shows the number of spring cubs, yearlings, 2-year-olds, and 3-year-old cubs seen in different years. The observations for the different years are not all comparable. During many of the early years, my studies were concentrated on other species which cut down on the observations of bears, and in some later years I was not in the park all summer so observations were less intensive. The years most comparable are from 1959 to 1969. One cannot assess cub losses very accurately from the table because some intact families may have been present but not seen.
Table 3. Minimum numbers of families of one, two,
and three cubs observed in Mt. McKinley Park, 1922-1970.

(click on image for a PDF version)
I obtain a crude assessment of cub loss by comparing numbers of spring cubs in each of the years 1959 to 1966 to the numbers of yearlings the following year, and to 2-year-olds 2 years later. Loss of spring cubs was 31%, whereas that of yearlings was 17%. These rates of loss are similar to those reported on the Alaskan peninsula (A. W. Stokes, pers. comm.) and in Yellowstone National Park (Craighead and Craighead 1967). Note the decline of families of three cubs with their age: 11 of spring cubs, 5 of yearlings, and 1 of 2-year-olds. This suggests that at least one cub from families of triplets is lost frequently. The increase of one-cub families with their age (21, 25, and 30 for spring, yearling, and 2-year-old cubs, respectively) also suggests loss from litters of two and three cubs.
Tracks and Trails
A few years ago, planners for the National Park Service suggested building trails for humans up some of the rivers and elsewhere in McKinley National Park. The grizzlies could have told them what park personnel learned later: that one can travel with relative ease over most of the park without trails.
Generally, grizzlies travel about at random. The few short pieces of bear trail that I have noted in the park have been along streams bordered by spruces, in stretches where bears frequently travel because of the terrain.
These bear trails, on the hard ground, remind me of Thoreau's trail at Walden in deep snow: ". . . For a week of even weather I took exactly the same number of steps, and of the same length, coming and going, stepping deliberately and with the precision of a pair of dividers in my own deep tracks. . . ." For some reason the bears tend to step in the same tracks until, over the years, a series of depressions is worn. In a short stretch of trail along the Teklanika River, where the water washed against the spruce-grown banks, the track depressions on the firm ground were worn an inch or more in depth, and were roughly 10 inches wide and a dozen inches in length. In these trails the front and hindfeet had stepped in the same depressions. The distance between them ranged from 23 to 30 inches. On a few occasions I have watched bears step in old tracks crossing snowfields, not missing a track. One bear followed a track in the snow as it walked to the top of a slope, stepping in the old tracks, then on reaching the top, he turned around and came down the slope carefully stepping in the tracks again.
In a slow, walking gait the hind foot may fall in the track of the forefoot or behind it, but usually in walking the hind foot registers ahead of the forefoot on the same side. In galloping, as we would expect, the tracks of the hind feet register ahead of the forefeet tracks in each set of four tracks. The pattern varies. Both hind feet may strike anterior to both forefeet, but sometimes one of the hind feet may be opposite the anterior forefoot track. In one set I noted that the tracks in each jump formed a diagonal line; the trail consisted of a series of these diagonal lines.
Bears show a great deal of variability in the way they move, but large males generally have a distinctive, ponderous walk, seemingly less flexible than females and younger individuals. Although grizzlies appear slow and somewhat ungainly when ambling along and feeding, they are well known to be capable of rapid bursts when galloping. On one occasion a galloping bear seemed to get most of its power from the front legs, the hind legs being brought forward without pushing. The forefeet landed well apart and the hind feet came down close together. But this was a subjective impression.
In spring and early summer, when most of the ground is free of snow, bear trails may be seen crossing the many snowfields lying in the draws of mountain slopes and in the hollows out on the rolling tundra. Some snowfields are crossed because they happen to lie in the line of travel, but I have the impression that bears prefer walking on the hard snow. On 3 June I observed a bear digging roots on a steep hillside. He started down the slope at a walk, catching himself at each step because of its steepness. I suggested to my companion that the bear could descend more easily if he would use the snowfield about forty yards to his right. A moment later he did just that and found travel easier. At first he sank in the snow with each step, but there was no jolting. Then, where the snowfield was firmer, he slid with hind legs trailing. The drift was softer lower down and he resumed wading, and where the slope became less steep, galloped in the snow as though he enjoyed the lark.
It also seems that grizzlies do not mind, and in fact apparently enjoy, a little sliding or "skiing," in this way resembling otters. Some slides are taken lying on one side. Slide marks on one slope showed that both a mother and cub had taken a rather long slide lying on their sides. On 17 May 1962, I watched a lone bear walk out on a snowfield, get mired, roll over to extricate himself, and then let himself slide while lying on his side. Near the base of the snowfield's slope he turned so as to face up the slope as he put on the brakes by digging in his claws. Coming to a stop, he continued to cross the snowfield on a lower contour line. His relaxed body and benign facial expression suggested he rather enjoyed the ride. Some snow trails consist of two parallel grooves coming, in some cases, directly down a slope. The bears making this kind of trail slide while standing in a skiing position. One bear appeared to have slid in this way for at least 200 feet (Fig. 12).
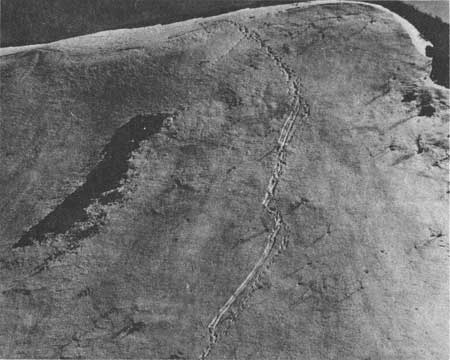
|
| Fig. 12. Bear trail descending steep snow slope. One of the bears did some skiing. |
One spring I watched a bear descending a snowfield into a deep ravine and beginning to slide on his feet. Being a discreet bear, he felt he was sliding too fast and turned by digging in his forepaws until he was facing up the slope. He stopped by braking with all four feet. He made a couple of jumps to the brink of the steepest part of the slope, then, before sliding, he turned so as to face up hill and in this position slid into the ravine out of my view.
The following day I saw this same bear wading in deep snow that had drifted into willow brush on a gentle slope. Instead of continuing the wading, he lay on his side and rolled like a barrel over the snow and willow tops. After rolling over four or five times, he reached the edge of the willow patch and started walking, but after a few steps, he lay down again to progress by the rolling technique, rolling over four or five more times. I expect that the scratching effect of the rolling was about as much incentive as was the ease of progress it contributed.
To what extent snow and dirt bother the feet by adhering between the toes is not known. But one day in October, when a light skiff of snow had fallen, three balls of mud, each about 1-1/2 inches in diameter, were picked up on a fresh trail. They had been pulled loose, apparently with the teeth, for a little hair was mixed with each one. Beyond the mud balls, a little blood was noted in the tracks. The mixture of snow and mud apparently had been just right for causing the mud to ball up.
The tracks of hind and front feet of the grizzly differ considerably. The main pad of the hindfoot is long; the claw marks extend only about 1 inch in front of the five toe-pad marks. The track of the forefoot consists of a short, broad pad mark posterior to five toe-pad marks. The tips of the long claws make marks about 2 inches in front of the five toe pads. If the track is deep, a small, rounded pad registers posterior to the main pad. Both front and hindfeet turn inward in walking.
Occasionally, the track of an individual may be distinctive. The main pad track of one hindfoot of a large male tapered toward the rear much more than it did in the other track. The difference was so obvious that this male's trail could be identified readily. A bear crippled on a front foot also left a distinctive track. The impression of the injured front foot was small and the marks of the unworn claws extended farther from the toes than in the normal foot.
On the river bars there are wet patches of mud or fine sand in which tracks show up well. However, a bear takes no pains to accommodate one with clear tracks. One day on the Toklat River bar, I saw a mother and yearling cross the river and move up along the bar. I followed to look for tracks, but even though there were many moist patches of mud scattered about, ideal for track impressions, they had all been avoided and not one track was seen. When one does find tracks either in mud or snow, they are often too ill-defined for accurate measurements. Sometimes a good front track may be found but no hindfoot track suitable for measurement. A track may be found that is satisfactory for measurement of width but not for length, so that one must examine several before finding measurable tracks. A series may vary in size because of slippage, depth of impression, rate of travel, or character of ground or snow. Hence, measurements obtained from a short trail may be rather miscellaneous in character and incomplete.
I have listed track measurements for both sexes and for cubs of different ages (Table 4). The claw marks are included in the measurements unless otherwise stated. In those for cubs there may be some variation due to differences in size between cubs of different families, and even in the same family. Tracks of the cubs in the different age groups tend, of course, to be larger in the fall than they are in the spring, due to the summer growth of cubs.
Table 4. Measurements of grizzly bear tracks. Single entries indicate measure of single tracks; others are a range from several tracks or, for cubs, from two litter mates.
| Front foot (inches) |
Hind foot (inches) |
Date | |||
| Width | Length | Width | Length | ||
| Adult female | 5—5-1/2 | 5 | 9 | ||
| Adult male | 6—6-3/4 | 6—6-1/2 | 10-3/4—12 | ||
| Spring cub | 3 | 3-5/8 | 25 July | ||
| 3 | 23 June | ||||
| 3-3/8—3-3/4 | 3-3/4 without claws | 6-1/4—7 | Aug—Sept. | ||
| Yearling cub | 4 | 7 | 10 June | ||
| 4 | 7 | 17 Aug. | |||
| 3-3/4—4 | 7-1/8—8 | 16 Sept. | |||
| 3-1/2 | 3 June | ||||
| Two-year-old cub | 4-1/4—4-1/2 | May | |||
| 4-1/2—5 | 4-1/2 | 9-5/8 | May | ||
| 4-1/4—4-1/2 | 7-3/4 | 4-1/4 | 7—7-1/2 | 3 June | |
Occasionally, one encounters a bear that is limping. He may have only a limp, or he may not use the foot in walking, or he may use it when walking but carry it when loping. Some bears have a permanent limp, while others may recover. In a later section dealing with grizzly—porcupine relationships, it is pointed out that a bear sometimes makes contact with a porcupine as indicated by quills sticking in the face or in a paw. A crippled bear that was killed had quills inside the crippled foot; apparently the quills had caused a permanent injury. A 2-year-old cub was stuck with quills, both in its face and one paw. Some other crippled bears will be described briefly in this section, for it would not be fair to the porcupine, or perhaps to the bear either, to list them all in the porcupine discussion.
On 13 June 1959, I saw a large, dark male with a severe limp of the left front foot. The elbow on the injured leg extended outward abnormally, and the foot turned inward excessively. The limp remained the same for the several weeks that the bear was seen. In 1963, this male grizzly was seen again, still with the same pronounced limp in the left front foot; obviously this bear had a permanent foot injury. One of two females mated to this male in 1959 limped on a hind leg. A round sore, the size of a dollar, could be seen just above the heel. When she galloped, the crippled foot was not used. She was first seen 17 June. On 1 July her foot had improved but she still carried it when she loped. On 4 July I saw her licking the sore spot; she limped a little but had improved, and by 10 July she seemed to have recovered. Over the years I observed several other bears, mainly cubs, with injured limbs. Of eight lame bears observed, six had injuries to a forefoot.
Injuries must be fairly rare and the causes of most is not known. Imprudent encounters with porcupines are certainly one source of such damage. Perhaps a cub occasionally is hurt slightly during over-exuberant play, especially with its mother, although I saw no evidence of this in several instances where females played roughly with their cubs. Sometimes a female, single-mindedly digging out a ground squirrel burrow on a rocky slope, will send large rocks flying below her, narrowly missing a cub. One spring cub was hit and rolled over by a rock about a foot in diameter in this situation, but did not seem to be injured.
Aside from their tracks, trails, and scats, "bear trees" are the other main sign of the presence of bears.
There is more than one kind of bear tree. When my brother and I were studying elk in northern Wyoming, we occasionally discovered trees with the bark torn loose near the base of the trunk. Species chiefly affected were the smooth-barked firs, but many lodgepole pines also were involved. The first time we encountered these trees we examined them closely and learned that the work was done by bears, apparently black bears. They were feeding on the cambium layer that carries the sap, seeking the raw syrup. The outer bark was bitten into and severed near the base of the trunk and the bark then pulled loose and stripped upward 3 to 4 feet or more so as to expose the cambium. Using the incisors, the cambium was scraped off, the close-set teeth leaving long, vertical, parallel grooves on the white, barkless trunk. The outer bark was pulled loose in several strips, occasionally around the entire trunk, so that when the bear was finished, these loose strips, attached 3 or 4 feet from the base, hung around the trunk like a grass skirt. Where such trees were found, there usually were several of them scattered in the area, sometimes 25 or 30. This suggested that once a bear tastes the "sweet" cambium delicacy he is reluctant to return to a more substantial diet. In some cases this feeding sign was both fresh and old. Perhaps the old sign was a reminder to a passing bear. In later years I also noted this feeding sign in the Olympic Mountains. We called them "bear trees."
When a black bear climbs an aspen tree, he leaves tracks. The claw marks heal over with scar tissue which remains for the life of the tree—a picturesque pattern registering a bygone event. These aspens are not at all uncommon in black bear—aspen country.
There is another kind of bear tree associated with both black and grizzly bears. These are trees situated so conveniently that they serve frequently as back scratchers. A lone tree along a trail or on the edge of a river bar is sure to be patronized often. Where many bears travel, even over a trail through a woods where numerous trees are available, there may be any number of bear trees showing signs of repeated use. In the Wood River country several miles east of McKinley National Park, I once followed for some distance a deeply worn bear trail through spruce woods bordering a high, perpendicular river bank. Bears passing up and down stream on that side of the river were somewhat hemmed in by the precipitous bank, sufficiently so that they generally used the trail. The traffic was so heavy that not only were the individual bear steps deeply worn, but much-used bear trees were closely spaced. I believe that the power of suggestion has given all bear travelers numerous itches, with the result that itching and rubbing on trees has increased through the years. I must add that all this bear sign on the crooked trail also increased my alertness, for at each turn I visualized a bear close enough for mutual embarrassment. Trails such as these, however, are rather scarce in the park, but along the Teklanika River there are short stretches of trails bordered by trees that show wear and have bear hairs embedded in pitch and lodged in the bark (Fig. 13).

|
| Fig. 13. A bear tree, used for rubbing. |
A bear tree may show scratches and tooth marks 6 or 7 feet up the trunk, and limbs have been broken off at 7-1/2 feet from the ground. In time the rubbing wears away patches of the bark. The ground is often worn smooth at the base of the tree. As the bear stands erect on hindfeet, with his back or stomach against the tree, or sits on haunches, he may bite randomly at the trunk, which, if the tree is slender, may produce two notches at different heights, roughly 3 and 5 feet. In a thorough scratching, the back, sides, rear, stomach, head, and neck are all massaged. Occasionally, the maneuvers suggest the latest "twist" dances as practiced by young people. One bear, standing on hind legs against a pole, raised and lowered herself, wriggling her body as part of the down movement to add to the effect.
In treeless country a large boulder is an excellent substitute for a tree. On Sable Pass I watched bears use a couple of poles that were lying on the ground, rolling on them so as to treat various parts of the anatomy. A few times I have seen a bear roll on the ground with much wriggling to get satisfaction, or to sit on haunches and rub his rear parts, which seemed to require excessive effort. Occasionally I have noted signs indicating that a bear had rubbed against a wooden bridge-railing and bitten off large slivers. Tall, stout willow brush is utilized sometimes. Frequently, bears rub on desirable sharp edges of log cabins and leave hairs. One morning, a large male rubbed against a log supporting the porch of Igloo Creek cabin and pushed it off its base. As they walk, bears often straddle brush or small spruce trees to scratch ventrally. No opportunities are overlooked.
Some of the literature suggests that trees are used by a bear to show other bears how high he can reach, hence how big he is, as a sort of warning to all to keep away from his domain. But in the first place, a grizzly does not lay claim to a domain. During the breeding season a bear tree might, I suppose, incidentally impart forcefully to a male the information that a desirable female passed that way. However, all observations indicate that the primary and conscious use of bear trees is for massaging.
When grizzlies encounter a pond in their travels, they may often wade in to lie down or take a swim, as though to cool off. They may also quench their thirst.
On 1 July 1940, what appeared to be a large male entered a small pond about 60 yards wide that lay in his line of travel. He splashed around a bit and swam across with body well submerged and nose pointed above the water. When he reached shore, he galloped for perhaps a half-mile.
On 8 September, 1939, I saw what appeared to be a young bear acting strangely. He hurried up a slope, then galloped down the ridge to Big Creek where he drank, then ran splashing down the middle of the stream. He was coming toward me, only 100 yards away, so I moved up the slope. When he came to my tracks, he reversed his direction, galloped up the stream, then over a ridge. He seemed to have enjoyed splashing his way down the stream, but the extreme exuberance was puzzling.
On 1 August, 1940, a female grizzly and spring cub walked along a small creek, feeding. It was a warm day and the female walked with open mouth and panted loudly. Three times she and the cub entered the creek, each time lying briefly in the water, obviously to cool off.
On 22 August 1950, Walter Weber and I saw a mother bear lying in a deep spot of Igloo Creek near Sable Pass, apparently cooling off.
On 15 September 1951, after a bear had rubbed his back on a pole, he sat down in a small puddle and appeared to wash his face with a paw.
On 24 July 1953, a mother and yearling were in a pond south of Cathedral Mountain. Part of the time the mother was completely submerged. They played a little in the water. The yearling soon went ashore but the mother remained in the cool water for several minutes.
On 28 May 1960, a large male entered a hollow and meandered around, attracted by the scent of meat. Earlier, two wolves were reported to have fed on something in this hollow and apparently had left only the scent of the food. After much frustration, the male walked into a pond in the hollow and lay with only his head and hump showing. A few times he submerged his head. Upon leaving the pond, he shook himself vigorously, did more searching for the source of the scent that was apparently strong in his nostrils, then re-entered the pond. When he left, he walked away without shaking, his wet hair flattened and dripping.
On 30 July 1962, the larger and more active of two yearlings entered a pond and swam and played for 3 or 4 minutes. The smaller yearling watched from the shore. He finally waded close to the shore, but only briefly. The mother continued to graze in the green hollow while the cubs were at the pond.
On 21 July 1953, a young bear spent several minutes wallowing in a pond, part of the time submerged. When he came out he seemed refreshed, approached a pole, bit into it, and then frisked away, galloping energetically. A bath often seemed to make the bears feel like romping.
On 27 August 1961, I watched two 2-year-old cubs that were in the process of separating from their mother straying off by themselves. After feeding for about 4 hours, they moved down to a small creek. One of them found a deep hole and waded in until the water covered all but his head. The other cub walked along in the middle of the stream for 30 yards and moved over to a green hollow to feed. It was a bright day, rather warm. I expect the cubs, in this rather casual manner, tarried in the water to cool off.
On 19 June 1955, a road worker watched a mother with spring cubs cross Igloo Creek, which still had some overflow ice protruding over the water in places. The spring cubs walked back and forth on shore, bawling, afraid to cross the rapid creek. One started across and was washed under the ice, but emerged farther downstream and managed to make the crossing. When my informant left, one of the cubs was still walking back and forth on shore, part of the time on his hind legs.
When streams are low, bears splash across without hesitation. But they recognize deep, fast water. On 14 June 1962, a lone bear, coming to a deep, rushing channel, stopped to ponder and walked along the edge circumspectly before entering. He was carried 50 yards downstream before reaching the other side.
The above notes are typical of the behavior often observed in the park. Along the coast of Alaska both black and brown bears spend much time in the water catching salmon, an activity not available to the McKinley National Park population of bears.
| <<< Previous | <<< Contents >>> | Next >>> |
chap2.htm
Last Updated: 06-Dec-2007